On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life
1
1950
... 自查尔斯·达尔文(Charles Darwin)和阿尔弗雷德·华莱士(Alfred Wallace)在19世纪50年代提出自然选择理论之后,生物类群如何适应环境一直是演化生物学的核心问题之一.自然选择理论认为种群中存在与表型相关的可遗传变异,其中有利于物种生存和繁殖的变异将被保留下来[1].在格雷戈尔·孟德尔(Gregor Mendel)发现的遗传规律及生物统计学和群体遗传学等理论基础上[2],“现代综合理论”(modern evolutionary synthesis)逐渐发展为理解表型适应性演化的重要理论框架[3]. ...
1
1971
... 自查尔斯·达尔文(Charles Darwin)和阿尔弗雷德·华莱士(Alfred Wallace)在19世纪50年代提出自然选择理论之后,生物类群如何适应环境一直是演化生物学的核心问题之一.自然选择理论认为种群中存在与表型相关的可遗传变异,其中有利于物种生存和繁殖的变异将被保留下来[1].在格雷戈尔·孟德尔(Gregor Mendel)发现的遗传规律及生物统计学和群体遗传学等理论基础上[2],“现代综合理论”(modern evolutionary synthesis)逐渐发展为理解表型适应性演化的重要理论框架[3]. ...
XV.: the correlation between relatives on the supposition of mendelian inheritance
1
1919
... 自查尔斯·达尔文(Charles Darwin)和阿尔弗雷德·华莱士(Alfred Wallace)在19世纪50年代提出自然选择理论之后,生物类群如何适应环境一直是演化生物学的核心问题之一.自然选择理论认为种群中存在与表型相关的可遗传变异,其中有利于物种生存和繁殖的变异将被保留下来[1].在格雷戈尔·孟德尔(Gregor Mendel)发现的遗传规律及生物统计学和群体遗传学等理论基础上[2],“现代综合理论”(modern evolutionary synthesis)逐渐发展为理解表型适应性演化的重要理论框架[3]. ...
Evolutionary developmental biology (evo-devo): past, present, and future
1
2012
... 在表型演化的遗传学研究基础上,演化发育生物学(evolutionary developmental biology)相关理论强调个体发育对于理解演化过程的重要意义[4].例如,同源异型基因在高度分歧的无脊椎动物和脊椎动物中发挥调控个体发育过程的功能.这些发育调控基因在不同生物类群中高度保守,同时又在同一类群中参与不同形态结构的发育过程,其编码区域可能受到强烈的功能限制[5].因此,演化发育理论强调复杂和模块化的调控网络的作用[6],认为其是基因功能的主要决定因素,且是形态变异和多样性的来源[7],为演化创新(evolutionary novelty)提供了理论基础.祖先中不存在的结构或性状可能来自新的基因调控网络[8],以及非编码区调控驱动了基因功能的创新[9]. ...
Hox gene variation and evolution
1
1998
... 在表型演化的遗传学研究基础上,演化发育生物学(evolutionary developmental biology)相关理论强调个体发育对于理解演化过程的重要意义[4].例如,同源异型基因在高度分歧的无脊椎动物和脊椎动物中发挥调控个体发育过程的功能.这些发育调控基因在不同生物类群中高度保守,同时又在同一类群中参与不同形态结构的发育过程,其编码区域可能受到强烈的功能限制[5].因此,演化发育理论强调复杂和模块化的调控网络的作用[6],认为其是基因功能的主要决定因素,且是形态变异和多样性的来源[7],为演化创新(evolutionary novelty)提供了理论基础.祖先中不存在的结构或性状可能来自新的基因调控网络[8],以及非编码区调控驱动了基因功能的创新[9]. ...
The developmental origin of novel complex morphological traits in Lepidoptera
1
2025
... 在表型演化的遗传学研究基础上,演化发育生物学(evolutionary developmental biology)相关理论强调个体发育对于理解演化过程的重要意义[4].例如,同源异型基因在高度分歧的无脊椎动物和脊椎动物中发挥调控个体发育过程的功能.这些发育调控基因在不同生物类群中高度保守,同时又在同一类群中参与不同形态结构的发育过程,其编码区域可能受到强烈的功能限制[5].因此,演化发育理论强调复杂和模块化的调控网络的作用[6],认为其是基因功能的主要决定因素,且是形态变异和多样性的来源[7],为演化创新(evolutionary novelty)提供了理论基础.祖先中不存在的结构或性状可能来自新的基因调控网络[8],以及非编码区调控驱动了基因功能的创新[9]. ...
Evo-devo and an expanding evolutionary synthesis: a genetic theory of morphological evolution
1
2008
... 在表型演化的遗传学研究基础上,演化发育生物学(evolutionary developmental biology)相关理论强调个体发育对于理解演化过程的重要意义[4].例如,同源异型基因在高度分歧的无脊椎动物和脊椎动物中发挥调控个体发育过程的功能.这些发育调控基因在不同生物类群中高度保守,同时又在同一类群中参与不同形态结构的发育过程,其编码区域可能受到强烈的功能限制[5].因此,演化发育理论强调复杂和模块化的调控网络的作用[6],认为其是基因功能的主要决定因素,且是形态变异和多样性的来源[7],为演化创新(evolutionary novelty)提供了理论基础.祖先中不存在的结构或性状可能来自新的基因调控网络[8],以及非编码区调控驱动了基因功能的创新[9]. ...
Evolutionary novelties
1
2010
... 在表型演化的遗传学研究基础上,演化发育生物学(evolutionary developmental biology)相关理论强调个体发育对于理解演化过程的重要意义[4].例如,同源异型基因在高度分歧的无脊椎动物和脊椎动物中发挥调控个体发育过程的功能.这些发育调控基因在不同生物类群中高度保守,同时又在同一类群中参与不同形态结构的发育过程,其编码区域可能受到强烈的功能限制[5].因此,演化发育理论强调复杂和模块化的调控网络的作用[6],认为其是基因功能的主要决定因素,且是形态变异和多样性的来源[7],为演化创新(evolutionary novelty)提供了理论基础.祖先中不存在的结构或性状可能来自新的基因调控网络[8],以及非编码区调控驱动了基因功能的创新[9]. ...
The noncoding RNA revolution-trashing old rules to forge new ones
1
2014
... 在表型演化的遗传学研究基础上,演化发育生物学(evolutionary developmental biology)相关理论强调个体发育对于理解演化过程的重要意义[4].例如,同源异型基因在高度分歧的无脊椎动物和脊椎动物中发挥调控个体发育过程的功能.这些发育调控基因在不同生物类群中高度保守,同时又在同一类群中参与不同形态结构的发育过程,其编码区域可能受到强烈的功能限制[5].因此,演化发育理论强调复杂和模块化的调控网络的作用[6],认为其是基因功能的主要决定因素,且是形态变异和多样性的来源[7],为演化创新(evolutionary novelty)提供了理论基础.祖先中不存在的结构或性状可能来自新的基因调控网络[8],以及非编码区调控驱动了基因功能的创新[9]. ...
Doublesex is a mimicry supergene
5
2014
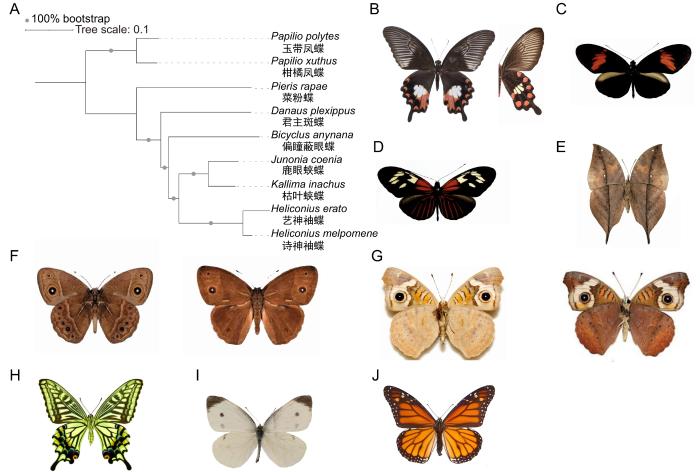
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
... [10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
...
玉带凤蝶(Papilio polytes) | 贝氏拟态[10] | | 美凤蝶(Papilio memnon) | 贝氏拟态[32,35] |
| 红斑美凤蝶(Papilio rumanzovia) | 贝氏拟态[35] |
| 果园美凤蝶(Papilio aegeus) | 贝氏拟态[35] |
optix ...
Evolution of diversity in warning color and mimicry: polymorphisms, shifting balance, and speciation
1
1999
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
The evolution and diversification of oakleaf butterflies
7
2022
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
... 枯叶蛱蝶属蝴蝶是著名的叶形伪装拟态动物,曾被英国博物学家与演化学家阿尔弗雷德·华莱士称为“蝴蝶伪装保护性的最美妙与最毋庸置疑的案例”[43].枯叶蛱蝶(Kallima inachus)处于静止状态时,展现出腹侧翅图案,模拟了黄棕色的枯萎叶片(图1E).其腹侧的拟叶图案还具有高度多态性,由形似枯叶主脉、次脉、叶柄和霉斑等的图案元件组合而成[44].这一独特的表型多态性也使其成为研究自然选择驱动下遗传多样性的良好体系.行为学实验表明,将幼年家鸡作为捕食者,经过训练的家鸡攻击枯叶蛱蝶与枯萎叶片的次数接近,证明了叶形伪装拟态表型的有效性[45].为了解析其翅图案的遗传和分子基础,研究人员基于核型实验、长读长测序技术和高通量染色体构象捕获(high-throughput chromosome conformation capture)测序技术,组装了枯叶蛱蝶染色体水平的高质量基因组[46];在后续研究中,又发现其叶形拟态表型由一个孟德尔基因座决定,进而定位到包含cortex基因在内的基因组区域[12].此外,对枯叶蛱蝶属内的跨物种研究提示,叶形拟态多态性的起源早于该属的物种分化[12].群体遗传学建模则表明,在枯叶蛱蝶的自然种群中,不同等位基因频率的观测数据符合负频率依赖选择驱动的平衡选择模型,表明该机制有助于维持叶形拟态多样性.上述研究提示:枯叶蛱蝶属的叶形伪装拟态表型是一个在物种分化过程中长期维持的、具有明确适应意义的平衡选择实例[12]. ...
... [12].群体遗传学建模则表明,在枯叶蛱蝶的自然种群中,不同等位基因频率的观测数据符合负频率依赖选择驱动的平衡选择模型,表明该机制有助于维持叶形拟态多样性.上述研究提示:枯叶蛱蝶属的叶形伪装拟态表型是一个在物种分化过程中长期维持的、具有明确适应意义的平衡选择实例[12]. ...
... [12]. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
... ,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
... | 枯叶蛱蝶属(Kallima) | 叶形拟态多样性[12] |
| 鹿眼蛱蝶(Junonia coenia) | 季节性表型可塑性[52] |
| 斑凤蝶(Papilio clytia) | 贝氏拟态[73] |
| 狐眼袖蝶(Heliconius numata) | 米勒拟态[74-75] |
| Distal-less | 转录因子 | 偏瞳蔽眼蝶(Bicyclus anynana) | 眼斑[48] |
未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
The evolutionary significance of dry and wet season forms in some tropical butterflies
1
1984
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
The effects of temperature and daylength on the Rosa polyphenism in the buckeye butterfly, Precis coenia (Lepidoptera: Nymphalidae)
1
1993
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
Colour constancy in the swallowtail butterfly Papilio Xuthus
2
2000
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
The genetic basis of a plant-insect coevolutionary key innovation
1
2007
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
Autumnal migration routes of the eastern population of the monarch butterfly (Danaus p. plexippus L.; Danaidae; Lepidoptera) in North America to the overwintering site in the Neovolcanic Plateau of Mexico
2
1978
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
... 君主斑蝶以其跨越北美的长距离迁飞[17]和鲜艳的橙色警戒色(图1J)闻名[18],但长期以来其遗传机制并不明确.相关研究通过对多种斑蝶属蝴蝶的基因组测序,揭示了君主斑蝶的物种起源、扩散历史以及长距离迁飞行为和警戒色的相关机制[68-69].全基因组系统发生树表明,君主斑蝶起源于北美,随后扩散至中美、南美、大西洋和太平洋地区,部分衍生种群逐渐丧失迁飞能力[69].为进一步识别与迁飞行为相关的变异,研究评估了种群分支指数(population branch statistic),并识别出约5.14 Mb的基因组区域与迁飞行为显著相关,其中共包括536个基因,基因本体(gene ontology)显示这些基因主要涉及神经发生和肌肉结构等功能[69].此外,相关研究也厘清了君主斑蝶警戒色的遗传基础.夏威夷地区存在一种白化突变型(nivosus),相较于野生型,其颜色由隐性等位基因控制.研究人员对白化型和野生型个体的基因组进行关联分析,显示肌球蛋白基因DPOGS20661与表型之间具有最强的关联水平,提示其翅图案表型可能是通过影响色素运输而改变[69]. ...
Heart poisons in the Monarch Butterfly: Some aposematic butterflies obtain protection from cardenolides present in their food plants
2
1968
... 蝶类昆虫具有多样的翅图案表型以及复杂的生理和行为特征,长期受到生物演化研究的关注.近年来,蝶类因其与外界环境之间复杂的相互作用机制成为生态、演化和行为学领域的研究热点,相关研究体系和研究方向不断拓展,涉及拟态策略、季节型与表型可塑性以及环境感知机制等多个方面.在蝶类拟态的相关研究中,形成了以玉带凤蝶(Papilio polytes)为代表的贝氏拟态(Batesian mimicry)研究体系[10]、以袖蝶属(Heliconius)诗神袖蝶(Heliconius melpomene)与艺神袖蝶(Heliconius erato)等构成的米勒拟态(Müllerian mimicry)研究体系[11],以及聚焦枯叶蛱蝶属(Kallima)[12]形成的叶形伪装拟态(leaf masquerade mimicry)研究体系,系统性研究物种规避捕食的不同拟态策略及其背后的遗传与演化基础.此外,蝶类对旱季与雨季环境的响应还表现出显著的季节型差异和表型可塑性[13-14].例如:鹿眼蛱蝶(Junonia coenia)与偏瞳蔽眼蝶(Bicyclus anynana)的翅图案展现出旱雨季差异,使其成为研究表型可塑性的经典体系.以上蝶类物种可用于研究生物个体在选择压力下对环境的动态适应能力.此外,蝶类体系还可用于探究生物与环境的复杂互动,例如:基于柑橘凤蝶(Papilio xuthus)研究其视觉感知模型[15]、基于菜粉蝶(Pieris rapae)研究蝶类与寄主植物相互作用[16],以及基于君主斑蝶(Danaus plexippus)研究超长距离迁飞行为[17]与警戒色[18]的协同演化等.上述研究体现了蝶类作为生态演化研究体系的优势和多样性. ...
... 君主斑蝶以其跨越北美的长距离迁飞[17]和鲜艳的橙色警戒色(图1J)闻名[18],但长期以来其遗传机制并不明确.相关研究通过对多种斑蝶属蝴蝶的基因组测序,揭示了君主斑蝶的物种起源、扩散历史以及长距离迁飞行为和警戒色的相关机制[68-69].全基因组系统发生树表明,君主斑蝶起源于北美,随后扩散至中美、南美、大西洋和太平洋地区,部分衍生种群逐渐丧失迁飞能力[69].为进一步识别与迁飞行为相关的变异,研究评估了种群分支指数(population branch statistic),并识别出约5.14 Mb的基因组区域与迁飞行为显著相关,其中共包括536个基因,基因本体(gene ontology)显示这些基因主要涉及神经发生和肌肉结构等功能[69].此外,相关研究也厘清了君主斑蝶警戒色的遗传基础.夏威夷地区存在一种白化突变型(nivosus),相较于野生型,其颜色由隐性等位基因控制.研究人员对白化型和野生型个体的基因组进行关联分析,显示肌球蛋白基因DPOGS20661与表型之间具有最强的关联水平,提示其翅图案表型可能是通过影响色素运输而改变[69]. ...
The genetics and evo-devo of butterfly wing patterns
1
2002
... 蝶类翅结构简单但图案模式高度多样[19],通常由一系列保守图案元件的模块化组合构成,被称为蛱蝶平面图(Nymphalid ground plan)[20-21].蛱蝶平面图由一组高度保守的表型元件组成,主要包括基部(basal)、中央(central)和边缘(border)3个“对称系统”(symmetry system)元件.中央和边缘对称系统的中线上常具有分离的色素点,分别形成了中室斑(discal spot)和眼点(border ocelli)元素.此外,在蛱蝶平面图框架下,翅脉是图案模式的另一个重要影响因素.翅脉将翅图案分隔为多个区域,并可能造成图案元件的中断,使同一对称系统中的不同区域能够以更大的自由度实现组合形式的改变,进一步增加翅图案表型多样性. ...
on the ground-plan of wing-pattern in nymphalids and certain other families of the rhopaloeerous Lepidoptera
1
21
... 蝶类翅结构简单但图案模式高度多样[19],通常由一系列保守图案元件的模块化组合构成,被称为蛱蝶平面图(Nymphalid ground plan)[20-21].蛱蝶平面图由一组高度保守的表型元件组成,主要包括基部(basal)、中央(central)和边缘(border)3个“对称系统”(symmetry system)元件.中央和边缘对称系统的中线上常具有分离的色素点,分别形成了中室斑(discal spot)和眼点(border ocelli)元素.此外,在蛱蝶平面图框架下,翅脉是图案模式的另一个重要影响因素.翅脉将翅图案分隔为多个区域,并可能造成图案元件的中断,使同一对称系统中的不同区域能够以更大的自由度实现组合形式的改变,进一步增加翅图案表型多样性. ...
1
1991
... 蝶类翅结构简单但图案模式高度多样[19],通常由一系列保守图案元件的模块化组合构成,被称为蛱蝶平面图(Nymphalid ground plan)[20-21].蛱蝶平面图由一组高度保守的表型元件组成,主要包括基部(basal)、中央(central)和边缘(border)3个“对称系统”(symmetry system)元件.中央和边缘对称系统的中线上常具有分离的色素点,分别形成了中室斑(discal spot)和眼点(border ocelli)元素.此外,在蛱蝶平面图框架下,翅脉是图案模式的另一个重要影响因素.翅脉将翅图案分隔为多个区域,并可能造成图案元件的中断,使同一对称系统中的不同区域能够以更大的自由度实现组合形式的改变,进一步增加翅图案表型多样性. ...
Transcription factors underlying wing margin color patterns and pupal cuticle markings in butterflies
1
2020
... 眼斑是鳞翅目昆虫翅图案组成中的重要元件之一,可在规避捕食者与性选择等过程中发挥功能,其位置、大小与色环构成的变化是研究翅图案形成机制的核心问题.在多个鳞翅目物种中,其翅图案形成相关的有沟(engrailed)、大斑(spalt major)和无远端同源框(Distal-less,Dll)等基因的表达位置均与成虫翅图案中的眼斑部位高度重合[22]. ...
Tympanal ears mediate male-male competition, courtship and mating success in Bicyclus anynana butterflies
1
2024
... 鳞翅目昆虫翅组织通常具有按顺序排布和覆盖的微小鳞片(wing scales),其形态结构从扁平叶状到毛状不等,是构成复杂翅图案的基本单位.具体而言,鳞片可能通过黑色素(melanins)、眼色素(ommo-chromes),以及蝶呤类色素(pteridines)等显色,上述化学色素吸收特定波段光线,形成相应的翅图案.鳞片也可能通过其特异的物理结构和光线入射角度呈现出具有金属光泽的结构色.色素色与结构色的组合使鳞翅目昆虫翅图案呈现出高度复杂性,在其伪装、防御以及性选择等过程中发挥重要功能.此外,蝶翅还能承担诸如求偶展示、警戒信号以及体温调节等多重功能[23].因此,蝶翅是在演化发育框架下,研究适应性性状演化的理想模型.近年来的蝶类演化发育研究聚焦于代表性蝶类翅花纹(图1),逐渐形成了表型─基因─机制的多层次研究范式. ...
1
1890
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
XXXII. Contributions to an insect fauna of the Amazon valley (Lepidoptera: Heliconidae)
1
1862
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
Super-genes and mimicry
1
1960
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
Diversity in mimicry: paradox or paradigm?
1
1998
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
The genetics of the mimetic butterfly Papilio memnon L
0
1968
Preferential predation of female butterflies and the evolution of batesian mimicry
1
1995
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
The diversity and evolution of Batesian mimicry in Papilio swallowtail butterflies
1
2009
... 拟态指的是在自然界长期生存压力下,一种生物模仿另一种生物,使其中一方受益或者双方都受益的现象[24].贝氏拟态由英国博物学家亨利·贝茨(Henry Bates)于1862年首次提出,是一种经典的生物拟态现象,描述的是无毒物种具有与有毒物种相似的形态,造成捕食者误判,导致无毒物种被捕食的概率降低[25].有观点认为,贝氏拟态由单个孟德尔位点控制,即多个紧密连锁基因构成的超基因位点[26].凤蝶属(Papilio)蝴蝶中广泛存在雌性特异性的贝氏拟态[27-29],其中雌性玉带凤蝶分为拟态和非拟态2类,拟态个体的翅图案(图1B)模拟有毒的红珠凤蝶(Pachliopta aristolochiae)[30].研究人员基于回交家系的图谱定位,将拟态位点定位至一个300 kb的基因组区域,其中包含昆虫的性别调控基因(doublesex,dsx),该基因在雌雄个体中分别具有不同的可变剪接形式[10].免疫荧光实验结果也表明,dsx基因在蝶翅发育过程中的空间表达模式与成虫拟态图案区域位置相同[10].总的来说,在玉带凤蝶中,dsx基因一方面维持其固有的性别分化功能,另一方面通过演化创新控制雌性特异的贝氏拟态多样性. ...
Genetic switch in UV response of mimicry-related pale-yellow colors in Batesian mimic butterfly, Papilio polytes
1
2021
... 有研究进一步显示,在玉带凤蝶翅盘发育过程中,dsx通过调控下游ebony与vermilion等色素合成基因的表达水平决定雌性特异拟态图案[31].除了dsx基因,该基因座内还包含广泛表达转录蛋白(ubiquitously expressed transcript)等基因,功能实验表明,以上基因共同调控玉带凤蝶拟态翅图案形成[32-34].对不同凤蝶属物种的跨物种比较则揭示了该基因座的演化历史.研究表明,多个凤蝶物种中的雌性特异性贝氏拟态均由dsx基因座决定,但物种间基因座结构和调控机制并不完全相同,提示其为拟态表型演化过程中的热点区域[35-36]. ...
Parallel evolution of Batesian mimicry supergene in two Papilio butterflies, P. polytes and P. memnon
2
2018
... 有研究进一步显示,在玉带凤蝶翅盘发育过程中,dsx通过调控下游ebony与vermilion等色素合成基因的表达水平决定雌性特异拟态图案[31].除了dsx基因,该基因座内还包含广泛表达转录蛋白(ubiquitously expressed transcript)等基因,功能实验表明,以上基因共同调控玉带凤蝶拟态翅图案形成[32-34].对不同凤蝶属物种的跨物种比较则揭示了该基因座的演化历史.研究表明,多个凤蝶物种中的雌性特异性贝氏拟态均由dsx基因座决定,但物种间基因座结构和调控机制并不完全相同,提示其为拟态表型演化过程中的热点区域[35-36]. ...
...
玉带凤蝶(Papilio polytes) | 贝氏拟态[10] | | 美凤蝶(Papilio memnon) | 贝氏拟态[32,35] |
| 红斑美凤蝶(Papilio rumanzovia) | 贝氏拟态[35] |
| 果园美凤蝶(Papilio aegeus) | 贝氏拟态[35] |
optix ...
Functional unit of supergene in female-limited Batesian mimicry of Papilio polytes
0
2023
Functional genetic elements of a butterfly mimicry supergene
1
2025
... 有研究进一步显示,在玉带凤蝶翅盘发育过程中,dsx通过调控下游ebony与vermilion等色素合成基因的表达水平决定雌性特异拟态图案[31].除了dsx基因,该基因座内还包含广泛表达转录蛋白(ubiquitously expressed transcript)等基因,功能实验表明,以上基因共同调控玉带凤蝶拟态翅图案形成[32-34].对不同凤蝶属物种的跨物种比较则揭示了该基因座的演化历史.研究表明,多个凤蝶物种中的雌性特异性贝氏拟态均由dsx基因座决定,但物种间基因座结构和调控机制并不完全相同,提示其为拟态表型演化过程中的热点区域[35-36]. ...
A shared genetic basis of mimicry across swallowtail butterflies points to ancestral co-option of doublesex
4
2020
... 有研究进一步显示,在玉带凤蝶翅盘发育过程中,dsx通过调控下游ebony与vermilion等色素合成基因的表达水平决定雌性特异拟态图案[31].除了dsx基因,该基因座内还包含广泛表达转录蛋白(ubiquitously expressed transcript)等基因,功能实验表明,以上基因共同调控玉带凤蝶拟态翅图案形成[32-34].对不同凤蝶属物种的跨物种比较则揭示了该基因座的演化历史.研究表明,多个凤蝶物种中的雌性特异性贝氏拟态均由dsx基因座决定,但物种间基因座结构和调控机制并不完全相同,提示其为拟态表型演化过程中的热点区域[35-36]. ...
... | 玉带凤蝶(Papilio polytes) | 贝氏拟态[10] |
| 美凤蝶(Papilio memnon) | 贝氏拟态[32,35] |
| 红斑美凤蝶(Papilio rumanzovia) | 贝氏拟态[35] |
| 果园美凤蝶(Papilio aegeus) | 贝氏拟态[35] |
optix ...
... [35] |
| 果园美凤蝶(Papilio aegeus) | 贝氏拟态[35] |
optix ...
... [35] |
optix ...
Genomic architecture and functional unit of mimicry supergene in female limited Batesian mimic Papilio butterflies
1
2022
... 有研究进一步显示,在玉带凤蝶翅盘发育过程中,dsx通过调控下游ebony与vermilion等色素合成基因的表达水平决定雌性特异拟态图案[31].除了dsx基因,该基因座内还包含广泛表达转录蛋白(ubiquitously expressed transcript)等基因,功能实验表明,以上基因共同调控玉带凤蝶拟态翅图案形成[32-34].对不同凤蝶属物种的跨物种比较则揭示了该基因座的演化历史.研究表明,多个凤蝶物种中的雌性特异性贝氏拟态均由dsx基因座决定,但物种间基因座结构和调控机制并不完全相同,提示其为拟态表型演化过程中的热点区域[35-36]. ...
Ituna and Thyridia: a remarkable case of mimicry in butterflies
1
1879
... 米勒拟态由德国生物学家弗里茨·米勒(Fritz Müller)命名[37],指的是有毒物种之间互相模拟以降低捕食者的学习成本,并提高捕食者学习效果的现象.袖蝶属蝴蝶广泛分布于中美洲和南美洲,其幼虫以有毒的西番莲属(Passiflora)植物为食[38],具有高度的物种和表型多样性[39].米勒拟态被认为是驱动袖蝶警戒色演化的主要力量之一[40].其中研究最充分的类群是诗神袖蝶(图1C)与艺神袖蝶(图1D),同一物种的不同地理亚种之间,翅图案存在分歧,而在同域分布的两个物种间翅图案趋同,相互构成米勒拟态[41].对袖蝶属蝴蝶拟态图案遗传机制的研究表明,转录因子optix是控制其翅图案变异的关键基因之一[42].功能研究显示,optix基因在蛹期翅盘中具有特定的时空表达模式,并对应于成虫中红色图案元件的分布[42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
Resource partitioning in passion vine butterflies
1
1978
... 米勒拟态由德国生物学家弗里茨·米勒(Fritz Müller)命名[37],指的是有毒物种之间互相模拟以降低捕食者的学习成本,并提高捕食者学习效果的现象.袖蝶属蝴蝶广泛分布于中美洲和南美洲,其幼虫以有毒的西番莲属(Passiflora)植物为食[38],具有高度的物种和表型多样性[39].米勒拟态被认为是驱动袖蝶警戒色演化的主要力量之一[40].其中研究最充分的类群是诗神袖蝶(图1C)与艺神袖蝶(图1D),同一物种的不同地理亚种之间,翅图案存在分歧,而在同域分布的两个物种间翅图案趋同,相互构成米勒拟态[41].对袖蝶属蝴蝶拟态图案遗传机制的研究表明,转录因子optix是控制其翅图案变异的关键基因之一[42].功能研究显示,optix基因在蛹期翅盘中具有特定的时空表达模式,并对应于成虫中红色图案元件的分布[42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
Why are there so many mimicry rings? Correlations between habitat, behaviour and mimicry in Heliconius butterflies
1
1995
... 米勒拟态由德国生物学家弗里茨·米勒(Fritz Müller)命名[37],指的是有毒物种之间互相模拟以降低捕食者的学习成本,并提高捕食者学习效果的现象.袖蝶属蝴蝶广泛分布于中美洲和南美洲,其幼虫以有毒的西番莲属(Passiflora)植物为食[38],具有高度的物种和表型多样性[39].米勒拟态被认为是驱动袖蝶警戒色演化的主要力量之一[40].其中研究最充分的类群是诗神袖蝶(图1C)与艺神袖蝶(图1D),同一物种的不同地理亚种之间,翅图案存在分歧,而在同域分布的两个物种间翅图案趋同,相互构成米勒拟态[41].对袖蝶属蝴蝶拟态图案遗传机制的研究表明,转录因子optix是控制其翅图案变异的关键基因之一[42].功能研究显示,optix基因在蛹期翅盘中具有特定的时空表达模式,并对应于成虫中红色图案元件的分布[42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
Studies of müllerian mimicry and its evolution in burnet moths and heliconid butterflies
1
1971
... 米勒拟态由德国生物学家弗里茨·米勒(Fritz Müller)命名[37],指的是有毒物种之间互相模拟以降低捕食者的学习成本,并提高捕食者学习效果的现象.袖蝶属蝴蝶广泛分布于中美洲和南美洲,其幼虫以有毒的西番莲属(Passiflora)植物为食[38],具有高度的物种和表型多样性[39].米勒拟态被认为是驱动袖蝶警戒色演化的主要力量之一[40].其中研究最充分的类群是诗神袖蝶(图1C)与艺神袖蝶(图1D),同一物种的不同地理亚种之间,翅图案存在分歧,而在同域分布的两个物种间翅图案趋同,相互构成米勒拟态[41].对袖蝶属蝴蝶拟态图案遗传机制的研究表明,转录因子optix是控制其翅图案变异的关键基因之一[42].功能研究显示,optix基因在蛹期翅盘中具有特定的时空表达模式,并对应于成虫中红色图案元件的分布[42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
Genomic hotspots of adaptation in butterfly wing pattern evolution
1
2008
... 米勒拟态由德国生物学家弗里茨·米勒(Fritz Müller)命名[37],指的是有毒物种之间互相模拟以降低捕食者的学习成本,并提高捕食者学习效果的现象.袖蝶属蝴蝶广泛分布于中美洲和南美洲,其幼虫以有毒的西番莲属(Passiflora)植物为食[38],具有高度的物种和表型多样性[39].米勒拟态被认为是驱动袖蝶警戒色演化的主要力量之一[40].其中研究最充分的类群是诗神袖蝶(图1C)与艺神袖蝶(图1D),同一物种的不同地理亚种之间,翅图案存在分歧,而在同域分布的两个物种间翅图案趋同,相互构成米勒拟态[41].对袖蝶属蝴蝶拟态图案遗传机制的研究表明,转录因子optix是控制其翅图案变异的关键基因之一[42].功能研究显示,optix基因在蛹期翅盘中具有特定的时空表达模式,并对应于成虫中红色图案元件的分布[42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
Optix drives the repeated convergent evolution of butterfly wing pattern mimicry
4
2011
... 米勒拟态由德国生物学家弗里茨·米勒(Fritz Müller)命名[37],指的是有毒物种之间互相模拟以降低捕食者的学习成本,并提高捕食者学习效果的现象.袖蝶属蝴蝶广泛分布于中美洲和南美洲,其幼虫以有毒的西番莲属(Passiflora)植物为食[38],具有高度的物种和表型多样性[39].米勒拟态被认为是驱动袖蝶警戒色演化的主要力量之一[40].其中研究最充分的类群是诗神袖蝶(图1C)与艺神袖蝶(图1D),同一物种的不同地理亚种之间,翅图案存在分歧,而在同域分布的两个物种间翅图案趋同,相互构成米勒拟态[41].对袖蝶属蝴蝶拟态图案遗传机制的研究表明,转录因子optix是控制其翅图案变异的关键基因之一[42].功能研究显示,optix基因在蛹期翅盘中具有特定的时空表达模式,并对应于成虫中红色图案元件的分布[42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
... [42].上述研究揭示了不同袖蝶物种通过同一基因的调控序列,独立演化出相似的翅图案表型,为理解拟态图案的趋同演化提供了分子证据与新视角.以上研究展示了现代演化发育生物学的研究范式,即通过整合分子生物学、遗传学和演化生物学的方法来解析复杂性状的演化机制. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
... | 袖蝶属(Heliconius) | 米勒拟态[42,71] |
| 银纹红袖蝶(Agraulis vanillae) | 橙色和红色翅图案[72] |
| 鹿眼蛱蝶(Junonia coenia) |
| 小红蛱蝶(Vanessa cardui) |
cortex ...
1
1889
... 枯叶蛱蝶属蝴蝶是著名的叶形伪装拟态动物,曾被英国博物学家与演化学家阿尔弗雷德·华莱士称为“蝴蝶伪装保护性的最美妙与最毋庸置疑的案例”[43].枯叶蛱蝶(Kallima inachus)处于静止状态时,展现出腹侧翅图案,模拟了黄棕色的枯萎叶片(图1E).其腹侧的拟叶图案还具有高度多态性,由形似枯叶主脉、次脉、叶柄和霉斑等的图案元件组合而成[44].这一独特的表型多态性也使其成为研究自然选择驱动下遗传多样性的良好体系.行为学实验表明,将幼年家鸡作为捕食者,经过训练的家鸡攻击枯叶蛱蝶与枯萎叶片的次数接近,证明了叶形伪装拟态表型的有效性[45].为了解析其翅图案的遗传和分子基础,研究人员基于核型实验、长读长测序技术和高通量染色体构象捕获(high-throughput chromosome conformation capture)测序技术,组装了枯叶蛱蝶染色体水平的高质量基因组[46];在后续研究中,又发现其叶形拟态表型由一个孟德尔基因座决定,进而定位到包含cortex基因在内的基因组区域[12].此外,对枯叶蛱蝶属内的跨物种研究提示,叶形拟态多态性的起源早于该属的物种分化[12].群体遗传学建模则表明,在枯叶蛱蝶的自然种群中,不同等位基因频率的观测数据符合负频率依赖选择驱动的平衡选择模型,表明该机制有助于维持叶形拟态多样性.上述研究提示:枯叶蛱蝶属的叶形伪装拟态表型是一个在物种分化过程中长期维持的、具有明确适应意义的平衡选择实例[12]. ...
Using comprehensive machine-learning models to classify complex morphological characters
1
2021
... 枯叶蛱蝶属蝴蝶是著名的叶形伪装拟态动物,曾被英国博物学家与演化学家阿尔弗雷德·华莱士称为“蝴蝶伪装保护性的最美妙与最毋庸置疑的案例”[43].枯叶蛱蝶(Kallima inachus)处于静止状态时,展现出腹侧翅图案,模拟了黄棕色的枯萎叶片(图1E).其腹侧的拟叶图案还具有高度多态性,由形似枯叶主脉、次脉、叶柄和霉斑等的图案元件组合而成[44].这一独特的表型多态性也使其成为研究自然选择驱动下遗传多样性的良好体系.行为学实验表明,将幼年家鸡作为捕食者,经过训练的家鸡攻击枯叶蛱蝶与枯萎叶片的次数接近,证明了叶形伪装拟态表型的有效性[45].为了解析其翅图案的遗传和分子基础,研究人员基于核型实验、长读长测序技术和高通量染色体构象捕获(high-throughput chromosome conformation capture)测序技术,组装了枯叶蛱蝶染色体水平的高质量基因组[46];在后续研究中,又发现其叶形拟态表型由一个孟德尔基因座决定,进而定位到包含cortex基因在内的基因组区域[12].此外,对枯叶蛱蝶属内的跨物种研究提示,叶形拟态多态性的起源早于该属的物种分化[12].群体遗传学建模则表明,在枯叶蛱蝶的自然种群中,不同等位基因频率的观测数据符合负频率依赖选择驱动的平衡选择模型,表明该机制有助于维持叶形拟态多样性.上述研究提示:枯叶蛱蝶属的叶形伪装拟态表型是一个在物种分化过程中长期维持的、具有明确适应意义的平衡选择实例[12]. ...
Nature's disguise: Empirical demonstration of dead-leaf masquerade in Kallima butterflies
1
2024
... 枯叶蛱蝶属蝴蝶是著名的叶形伪装拟态动物,曾被英国博物学家与演化学家阿尔弗雷德·华莱士称为“蝴蝶伪装保护性的最美妙与最毋庸置疑的案例”[43].枯叶蛱蝶(Kallima inachus)处于静止状态时,展现出腹侧翅图案,模拟了黄棕色的枯萎叶片(图1E).其腹侧的拟叶图案还具有高度多态性,由形似枯叶主脉、次脉、叶柄和霉斑等的图案元件组合而成[44].这一独特的表型多态性也使其成为研究自然选择驱动下遗传多样性的良好体系.行为学实验表明,将幼年家鸡作为捕食者,经过训练的家鸡攻击枯叶蛱蝶与枯萎叶片的次数接近,证明了叶形伪装拟态表型的有效性[45].为了解析其翅图案的遗传和分子基础,研究人员基于核型实验、长读长测序技术和高通量染色体构象捕获(high-throughput chromosome conformation capture)测序技术,组装了枯叶蛱蝶染色体水平的高质量基因组[46];在后续研究中,又发现其叶形拟态表型由一个孟德尔基因座决定,进而定位到包含cortex基因在内的基因组区域[12].此外,对枯叶蛱蝶属内的跨物种研究提示,叶形拟态多态性的起源早于该属的物种分化[12].群体遗传学建模则表明,在枯叶蛱蝶的自然种群中,不同等位基因频率的观测数据符合负频率依赖选择驱动的平衡选择模型,表明该机制有助于维持叶形拟态多样性.上述研究提示:枯叶蛱蝶属的叶形伪装拟态表型是一个在物种分化过程中长期维持的、具有明确适应意义的平衡选择实例[12]. ...
Chromosome-level reference genome assembly and gene editing of the dead-leaf butterfly Kallima inachus
1
2020
... 枯叶蛱蝶属蝴蝶是著名的叶形伪装拟态动物,曾被英国博物学家与演化学家阿尔弗雷德·华莱士称为“蝴蝶伪装保护性的最美妙与最毋庸置疑的案例”[43].枯叶蛱蝶(Kallima inachus)处于静止状态时,展现出腹侧翅图案,模拟了黄棕色的枯萎叶片(图1E).其腹侧的拟叶图案还具有高度多态性,由形似枯叶主脉、次脉、叶柄和霉斑等的图案元件组合而成[44].这一独特的表型多态性也使其成为研究自然选择驱动下遗传多样性的良好体系.行为学实验表明,将幼年家鸡作为捕食者,经过训练的家鸡攻击枯叶蛱蝶与枯萎叶片的次数接近,证明了叶形伪装拟态表型的有效性[45].为了解析其翅图案的遗传和分子基础,研究人员基于核型实验、长读长测序技术和高通量染色体构象捕获(high-throughput chromosome conformation capture)测序技术,组装了枯叶蛱蝶染色体水平的高质量基因组[46];在后续研究中,又发现其叶形拟态表型由一个孟德尔基因座决定,进而定位到包含cortex基因在内的基因组区域[12].此外,对枯叶蛱蝶属内的跨物种研究提示,叶形拟态多态性的起源早于该属的物种分化[12].群体遗传学建模则表明,在枯叶蛱蝶的自然种群中,不同等位基因频率的观测数据符合负频率依赖选择驱动的平衡选择模型,表明该机制有助于维持叶形拟态多样性.上述研究提示:枯叶蛱蝶属的叶形伪装拟态表型是一个在物种分化过程中长期维持的、具有明确适应意义的平衡选择实例[12]. ...
Seasonal plasticity: how do butterfly wing pattern traits evolve environmental responsiveness?
1
2021
... 表型可塑性指同一物种中相同基因型的个体可以响应不同的环境信号而产生不同的表型[47].偏瞳蔽眼蝶是一种产于非洲的蛱蝶科蝴蝶,广泛分布于撒哈拉以南地区,其翅图案在雨季表现为黄褐色,眼斑大而明显(图1F);在旱季则是暗褐色,几乎无眼斑[48].为了解析偏瞳蔽眼蝶翅图案旱雨季差异的相关机制,研究人员基于免疫荧光技术检测了3种与眼斑相关的蛋白Notch、Engrailed和Distal-less在旱雨季型眼斑中的表达模式.其结果表明,Notch和Engrailed在旱季较雨季更早地在眼斑中心表达,这提示眼斑相关基因表达时间的差异参与调控了翅图案的季节型分化[49].研究者进一步通过在2种饲养环境下采集4个发育阶段的偏瞳蔽眼蝶幼虫翅盘样本,并利用转录组测序技术展示了翅发育过程中的基因表达模式.研究结果显示,相较于环境因素,发育阶段转变对基因表达水平、可变剪接和miRNA介导的基因沉默具有更强烈的影响[50].同时,研究结果也揭示了daughterless基因的可变剪接形式在2种季节型之间存在差异,这提示环境因素可能通过转录后调控机制影响蝶翅发育和表型可塑性[50]. ...
Development, plasticity and evolution of butterfly eyespot patterns
2
1996
... 表型可塑性指同一物种中相同基因型的个体可以响应不同的环境信号而产生不同的表型[47].偏瞳蔽眼蝶是一种产于非洲的蛱蝶科蝴蝶,广泛分布于撒哈拉以南地区,其翅图案在雨季表现为黄褐色,眼斑大而明显(图1F);在旱季则是暗褐色,几乎无眼斑[48].为了解析偏瞳蔽眼蝶翅图案旱雨季差异的相关机制,研究人员基于免疫荧光技术检测了3种与眼斑相关的蛋白Notch、Engrailed和Distal-less在旱雨季型眼斑中的表达模式.其结果表明,Notch和Engrailed在旱季较雨季更早地在眼斑中心表达,这提示眼斑相关基因表达时间的差异参与调控了翅图案的季节型分化[49].研究者进一步通过在2种饲养环境下采集4个发育阶段的偏瞳蔽眼蝶幼虫翅盘样本,并利用转录组测序技术展示了翅发育过程中的基因表达模式.研究结果显示,相较于环境因素,发育阶段转变对基因表达水平、可变剪接和miRNA介导的基因沉默具有更强烈的影响[50].同时,研究结果也揭示了daughterless基因的可变剪接形式在2种季节型之间存在差异,这提示环境因素可能通过转录后调控机制影响蝶翅发育和表型可塑性[50]. ...
... | 枯叶蛱蝶属(Kallima) | 叶形拟态多样性[12] |
| 鹿眼蛱蝶(Junonia coenia) | 季节性表型可塑性[52] |
| 斑凤蝶(Papilio clytia) | 贝氏拟态[73] |
| 狐眼袖蝶(Heliconius numata) | 米勒拟态[74-75] |
| Distal-less | 转录因子 | 偏瞳蔽眼蝶(Bicyclus anynana) | 眼斑[48] |
未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
Temporal gene expression variation associated with eyespot size plasticity in Bicyclus anynana
2
2013
... 表型可塑性指同一物种中相同基因型的个体可以响应不同的环境信号而产生不同的表型[47].偏瞳蔽眼蝶是一种产于非洲的蛱蝶科蝴蝶,广泛分布于撒哈拉以南地区,其翅图案在雨季表现为黄褐色,眼斑大而明显(图1F);在旱季则是暗褐色,几乎无眼斑[48].为了解析偏瞳蔽眼蝶翅图案旱雨季差异的相关机制,研究人员基于免疫荧光技术检测了3种与眼斑相关的蛋白Notch、Engrailed和Distal-less在旱雨季型眼斑中的表达模式.其结果表明,Notch和Engrailed在旱季较雨季更早地在眼斑中心表达,这提示眼斑相关基因表达时间的差异参与调控了翅图案的季节型分化[49].研究者进一步通过在2种饲养环境下采集4个发育阶段的偏瞳蔽眼蝶幼虫翅盘样本,并利用转录组测序技术展示了翅发育过程中的基因表达模式.研究结果显示,相较于环境因素,发育阶段转变对基因表达水平、可变剪接和miRNA介导的基因沉默具有更强烈的影响[50].同时,研究结果也揭示了daughterless基因的可变剪接形式在2种季节型之间存在差异,这提示环境因素可能通过转录后调控机制影响蝶翅发育和表型可塑性[50]. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
A transcriptomic atlas underlying developmental plasticity of seasonal forms of Bicyclus anynana butterflies
3
2022
... 表型可塑性指同一物种中相同基因型的个体可以响应不同的环境信号而产生不同的表型[47].偏瞳蔽眼蝶是一种产于非洲的蛱蝶科蝴蝶,广泛分布于撒哈拉以南地区,其翅图案在雨季表现为黄褐色,眼斑大而明显(图1F);在旱季则是暗褐色,几乎无眼斑[48].为了解析偏瞳蔽眼蝶翅图案旱雨季差异的相关机制,研究人员基于免疫荧光技术检测了3种与眼斑相关的蛋白Notch、Engrailed和Distal-less在旱雨季型眼斑中的表达模式.其结果表明,Notch和Engrailed在旱季较雨季更早地在眼斑中心表达,这提示眼斑相关基因表达时间的差异参与调控了翅图案的季节型分化[49].研究者进一步通过在2种饲养环境下采集4个发育阶段的偏瞳蔽眼蝶幼虫翅盘样本,并利用转录组测序技术展示了翅发育过程中的基因表达模式.研究结果显示,相较于环境因素,发育阶段转变对基因表达水平、可变剪接和miRNA介导的基因沉默具有更强烈的影响[50].同时,研究结果也揭示了daughterless基因的可变剪接形式在2种季节型之间存在差异,这提示环境因素可能通过转录后调控机制影响蝶翅发育和表型可塑性[50]. ...
... [50]. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
Hormonal control of a seasonal polyphenism in Precis coenia (Lepidoptera: Nymphalidae)
1
1995
... 鹿眼蛱蝶是一种广泛分布于北美的蛱蝶科蝴蝶,其翅图案在雨季表现为浅褐色(图1G),而在旱季则表现为暗红色[51].研究人员通过控制光周期和温度等环境因素,培养并建立了对环境响应能力不同的鹿眼蛱蝶实验室品系,并利用多组学研究方法探究了翅图案多态性的相关遗传机制[52].研究鉴定到3个参与可塑性调控的cortex、trehalase和herfst候选基因.基于成簇的规律间隔的短回文重复序列(clustered regularly interspaced short palindromic repeats,CRISPR)/CRISPR相关蛋白核酸酶9(CRISPR-associated nuclease 9,Cas9)基因编辑技术进行的功能验证表明,上述3个基因的功能缺失突变均可影响鹿眼蛱蝶翅颜色和眼斑大小[52],提示多个基因共同参与了表型调控.对相关基因调控元件的研究则凸显了顺式作用元件变异在调节季节型发育可塑性中的关键作用. ...
Genomic architecture of a genetically assimilated seasonal color pattern
5
2020
... 鹿眼蛱蝶是一种广泛分布于北美的蛱蝶科蝴蝶,其翅图案在雨季表现为浅褐色(图1G),而在旱季则表现为暗红色[51].研究人员通过控制光周期和温度等环境因素,培养并建立了对环境响应能力不同的鹿眼蛱蝶实验室品系,并利用多组学研究方法探究了翅图案多态性的相关遗传机制[52].研究鉴定到3个参与可塑性调控的cortex、trehalase和herfst候选基因.基于成簇的规律间隔的短回文重复序列(clustered regularly interspaced short palindromic repeats,CRISPR)/CRISPR相关蛋白核酸酶9(CRISPR-associated nuclease 9,Cas9)基因编辑技术进行的功能验证表明,上述3个基因的功能缺失突变均可影响鹿眼蛱蝶翅颜色和眼斑大小[52],提示多个基因共同参与了表型调控.对相关基因调控元件的研究则凸显了顺式作用元件变异在调节季节型发育可塑性中的关键作用. ...
... [52],提示多个基因共同参与了表型调控.对相关基因调控元件的研究则凸显了顺式作用元件变异在调节季节型发育可塑性中的关键作用. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
... ,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
...
枯叶蛱蝶属(Kallima) | 叶形拟态多样性[12] | | 鹿眼蛱蝶(Junonia coenia) | 季节性表型可塑性[52] |
| 斑凤蝶(Papilio clytia) | 贝氏拟态[73] |
| 狐眼袖蝶(Heliconius numata) | 米勒拟态[74-75] |
| Distal-less | 转录因子 | 偏瞳蔽眼蝶(Bicyclus anynana) | 眼斑[48] |
未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
The ivory lncRNA regulates seasonal color patterns in buckeye butterflies
2
2024
... 近期对鹿眼蛱蝶的研究进一步揭示了长链非编码RNA(long non-coding RNA)ivory在调控其季节型中的关键作用.ivory位于cortex基因座,通过整合分析该基因座的基因组结构、染色质开放状态及关键转录因子结合模式,揭示了其在翅图案模式调控中的核心作用,且在不同蝴蝶物种间高度保守[53].通过基因编辑技术突变ivory的顺式调控元件,得到了与季节型转变相似的表型变异,进一步验证了ivory在鹿眼蛱蝶环境适应和表型可塑性中的关键作用[53]. ...
... [53]. ...
Unconventional lift-generating mechanisms in free-flying butterflies
1
2002
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Butterfly host plant choice in the face of possible confusion
1
2000
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Understanding and misunderstanding the migration of the monarch butterfly (Nymphalidae) in North America: 1857-1995
1
1995
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Spectral organization of the eye of a butterfly, Papilio
1
2003
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Colour vision of the foraging swallowtail butterfly Papilio xuthus
1
1999
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Simultaneous color contrast in the foraging swallowtail butterfly, Papilio xuthus
1
2008
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Motion vision is independent of color in Drosophila
1
2008
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Monopolatic motion vision in the butterfly Papilio xuthus
1
2019
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
Motion-sensitive neurons activated by chromatic contrast in a butterfly visual system
2
2022
... 蝶类对环境信号的精准感知,表现出多种生态适应性行为,如飞行过程中的动态适应[54]、对寄主植物的识别与选择[55]、对季节与气候变化的迁飞响应[56]等,这些行为在其生命周期与演化适应中具有关键意义.例如,柑橘凤蝶(图1H)具有高度发达的视觉系统,是研究昆虫色彩识别的经典模式.柑橘凤蝶的复眼有6种光谱受体(紫外线、紫色、蓝色、绿色、红色以及宽波段),其复眼由多个小眼组成,每个小眼均包括9种感光细胞[57].行为学实验表明,柑橘凤蝶经过训练后可以正确识别多种颜色[15,58],其色彩识别方式与人类视觉中的对比色相同[59].在运动视觉方面,大部分昆虫[例如果蝇(Drosophila)]在飞行中依靠亮度变化而非色彩识别[60];然而,柑橘凤蝶依靠其复杂视觉系统感知光的线性偏振以调整飞行中的运动模式[61].电生理学研究表明,柑橘凤蝶具有特殊的视神经元,促进其在飞行过程中识别较小的移动目标[62].以上研究揭示了柑橘凤蝶运动视觉的高度复杂性,使其成为研究昆虫视觉机制和运动行为决策的重要生物模型. ...
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
The chemical diversity and distribution of glucosinolates and isothiocyanates among plants
1
2001
... 蝶类与其寄主植物之间还存在复杂的相互作用关系,菜粉蝶(图1I)分布广泛,是研究蝶类与寄主植物相互作用的典型研究体系.其寄主十字花科(Brassicaceae)植物被昆虫取食后会分泌有毒的次生代谢产物,如硫代葡萄糖苷[63].然而,菜粉蝶幼虫取食十字花科植物后,其肠道内一种特异性的酶会将硫代葡萄糖苷分解为腈类物质,并随幼虫粪便排出[64].此外,其寄主植物受到菜粉蝶幼虫取食后会通过茉莉酸信号通路释放挥发性有机化合物,以吸引寄生蜂等菜粉蝶天敌[65-67],从而形成三方相互作用的防御机制. ...
Successful herbivore attack due to metabolic diversion of a plant chemical defense
1
2004
... 蝶类与其寄主植物之间还存在复杂的相互作用关系,菜粉蝶(图1I)分布广泛,是研究蝶类与寄主植物相互作用的典型研究体系.其寄主十字花科(Brassicaceae)植物被昆虫取食后会分泌有毒的次生代谢产物,如硫代葡萄糖苷[63].然而,菜粉蝶幼虫取食十字花科植物后,其肠道内一种特异性的酶会将硫代葡萄糖苷分解为腈类物质,并随幼虫粪便排出[64].此外,其寄主植物受到菜粉蝶幼虫取食后会通过茉莉酸信号通路释放挥发性有机化合物,以吸引寄生蜂等菜粉蝶天敌[65-67],从而形成三方相互作用的防御机制. ...
GC-EAG-analysis of volatiles from Brussels sprouts plants damaged by two species of Pieris caterpillars: olfactory receptive range of a specialist and a generalist parasitoid wasp species
1
2002
... 蝶类与其寄主植物之间还存在复杂的相互作用关系,菜粉蝶(图1I)分布广泛,是研究蝶类与寄主植物相互作用的典型研究体系.其寄主十字花科(Brassicaceae)植物被昆虫取食后会分泌有毒的次生代谢产物,如硫代葡萄糖苷[63].然而,菜粉蝶幼虫取食十字花科植物后,其肠道内一种特异性的酶会将硫代葡萄糖苷分解为腈类物质,并随幼虫粪便排出[64].此外,其寄主植物受到菜粉蝶幼虫取食后会通过茉莉酸信号通路释放挥发性有机化合物,以吸引寄生蜂等菜粉蝶天敌[65-67],从而形成三方相互作用的防御机制. ...
Impact of herbivore-induced plant volatiles on parasitoid foraging success: a spatial simulation of the Cotesia rubecula, Pieris rapae, and Brassica oleracea system
0
2008
Jasmonic acid-induced volatiles of Brassica oleracea attract parasitoids: effects of time and dose, and comparison with induction by herbivores
1
2009
... 蝶类与其寄主植物之间还存在复杂的相互作用关系,菜粉蝶(图1I)分布广泛,是研究蝶类与寄主植物相互作用的典型研究体系.其寄主十字花科(Brassicaceae)植物被昆虫取食后会分泌有毒的次生代谢产物,如硫代葡萄糖苷[63].然而,菜粉蝶幼虫取食十字花科植物后,其肠道内一种特异性的酶会将硫代葡萄糖苷分解为腈类物质,并随幼虫粪便排出[64].此外,其寄主植物受到菜粉蝶幼虫取食后会通过茉莉酸信号通路释放挥发性有机化合物,以吸引寄生蜂等菜粉蝶天敌[65-67],从而形成三方相互作用的防御机制. ...
The monarch butterfly genome yields insights into long-distance migration
1
2011
... 君主斑蝶以其跨越北美的长距离迁飞[17]和鲜艳的橙色警戒色(图1J)闻名[18],但长期以来其遗传机制并不明确.相关研究通过对多种斑蝶属蝴蝶的基因组测序,揭示了君主斑蝶的物种起源、扩散历史以及长距离迁飞行为和警戒色的相关机制[68-69].全基因组系统发生树表明,君主斑蝶起源于北美,随后扩散至中美、南美、大西洋和太平洋地区,部分衍生种群逐渐丧失迁飞能力[69].为进一步识别与迁飞行为相关的变异,研究评估了种群分支指数(population branch statistic),并识别出约5.14 Mb的基因组区域与迁飞行为显著相关,其中共包括536个基因,基因本体(gene ontology)显示这些基因主要涉及神经发生和肌肉结构等功能[69].此外,相关研究也厘清了君主斑蝶警戒色的遗传基础.夏威夷地区存在一种白化突变型(nivosus),相较于野生型,其颜色由隐性等位基因控制.研究人员对白化型和野生型个体的基因组进行关联分析,显示肌球蛋白基因DPOGS20661与表型之间具有最强的关联水平,提示其翅图案表型可能是通过影响色素运输而改变[69]. ...
The genetics of monarch butterfly migration and warning colouration
4
2014
... 君主斑蝶以其跨越北美的长距离迁飞[17]和鲜艳的橙色警戒色(图1J)闻名[18],但长期以来其遗传机制并不明确.相关研究通过对多种斑蝶属蝴蝶的基因组测序,揭示了君主斑蝶的物种起源、扩散历史以及长距离迁飞行为和警戒色的相关机制[68-69].全基因组系统发生树表明,君主斑蝶起源于北美,随后扩散至中美、南美、大西洋和太平洋地区,部分衍生种群逐渐丧失迁飞能力[69].为进一步识别与迁飞行为相关的变异,研究评估了种群分支指数(population branch statistic),并识别出约5.14 Mb的基因组区域与迁飞行为显著相关,其中共包括536个基因,基因本体(gene ontology)显示这些基因主要涉及神经发生和肌肉结构等功能[69].此外,相关研究也厘清了君主斑蝶警戒色的遗传基础.夏威夷地区存在一种白化突变型(nivosus),相较于野生型,其颜色由隐性等位基因控制.研究人员对白化型和野生型个体的基因组进行关联分析,显示肌球蛋白基因DPOGS20661与表型之间具有最强的关联水平,提示其翅图案表型可能是通过影响色素运输而改变[69]. ...
... [69].为进一步识别与迁飞行为相关的变异,研究评估了种群分支指数(population branch statistic),并识别出约5.14 Mb的基因组区域与迁飞行为显著相关,其中共包括536个基因,基因本体(gene ontology)显示这些基因主要涉及神经发生和肌肉结构等功能[69].此外,相关研究也厘清了君主斑蝶警戒色的遗传基础.夏威夷地区存在一种白化突变型(nivosus),相较于野生型,其颜色由隐性等位基因控制.研究人员对白化型和野生型个体的基因组进行关联分析,显示肌球蛋白基因DPOGS20661与表型之间具有最强的关联水平,提示其翅图案表型可能是通过影响色素运输而改变[69]. ...
... [69].此外,相关研究也厘清了君主斑蝶警戒色的遗传基础.夏威夷地区存在一种白化突变型(nivosus),相较于野生型,其颜色由隐性等位基因控制.研究人员对白化型和野生型个体的基因组进行关联分析,显示肌球蛋白基因DPOGS20661与表型之间具有最强的关联水平,提示其翅图案表型可能是通过影响色素运输而改变[69]. ...
... [69]. ...
The industrial melanism mutation in British peppered moths is a transposable element
1
2016
... 综上所述,通过构建代表性蝶类演化研究体系并开展系统性研究,提示蝶类在翅表型、行为与生理层面展现出高度的适应性与多样性.蝶类物种在自然选择的压力下通过不同拟态与伪装策略以逃避捕食者捕捉,其中与拟态相关的翅图案花纹获得了较多关注,多个关键基因(表1)在不同物种翅图案花纹的演化发育中反复发挥着重要作用[12,52,70].在部分物种中,例如上文提及的贝氏拟态、米勒拟态以及叶形伪装拟态的蝶类研究体系中,发现复杂的蝶翅图案性状由相对简单的遗传机制决定,帮助其适应复杂的局部环境[10,12,42].鹿眼蛱蝶与偏瞳蔽眼蝶是用于研究旱雨季型与表型可塑性的经典研究体系,近期的研究发现在遗传差异之外[49,52],其相关分子机制可能还涉及生理和表观遗传,而环境因素与遗传背景通过复杂的基因调控网络以影响蝶类的翅图案模式[50].除以上与翅图案相关的演化研究外,蝶类体系还被广泛应用于其他生物学和生态学研究,例如,柑橘凤蝶因其动态视觉而成为研究昆虫视觉的典型体系[62],涵盖了生态行为学与神经生物学等多个层面;围绕菜粉蝶与其寄主植物之间相互作用为核心的研究体系,构建了有关生物相互作用以及功能适应的重要理论框架;以君主斑蝶为研究体系,解析了其迁飞种群演化历史以及飞行代谢响应等相关机制. ...
Multiple recent co-options of Optix associated with novel traits in adaptive butterfly wing radiations
1
2014
...
袖蝶属(Heliconius) | 米勒拟态[42,71] | | 银纹红袖蝶(Agraulis vanillae) | 橙色和红色翅图案[72] |
| 鹿眼蛱蝶(Junonia coenia) |
| 小红蛱蝶(Vanessa cardui) |
cortex ...
Single master regulatory gene coordinates the evolution and development of butterfly color and iridescence
1
2017
| 袖蝶属(Heliconius) | 米勒拟态[42,71] |
| 银纹红袖蝶(Agraulis vanillae) | 橙色和红色翅图案[72] |
| 鹿眼蛱蝶(Junonia coenia) |
| 小红蛱蝶(Vanessa cardui) |
cortex ...
Butterfly mimicry polymorphisms highlight phylogenetic limits of gene reuse in the evolution of diverse adaptations
1
2019
| 枯叶蛱蝶属(Kallima) | 叶形拟态多样性[12] |
| 鹿眼蛱蝶(Junonia coenia) | 季节性表型可塑性[52] |
| 斑凤蝶(Papilio clytia) | 贝氏拟态[73] |
| 狐眼袖蝶(Heliconius numata) | 米勒拟态[74-75] |
| Distal-less | 转录因子 | 偏瞳蔽眼蝶(Bicyclus anynana) | 眼斑[48] |
未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
A major gene controls mimicry and crypsis in butterflies and moths
1
2016
...
枯叶蛱蝶属(Kallima) | 叶形拟态多样性[12] | | 鹿眼蛱蝶(Junonia coenia) | 季节性表型可塑性[52] |
| 斑凤蝶(Papilio clytia) | 贝氏拟态[73] |
| 狐眼袖蝶(Heliconius numata) | 米勒拟态[74-75] |
| Distal-less | 转录因子 | 偏瞳蔽眼蝶(Bicyclus anynana) | 眼斑[48] |
未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
Chromosomal rearrangements maintain a polymorphic supergene controlling butterfly mimicry
1
2011
...
枯叶蛱蝶属(Kallima) | 叶形拟态多样性[12] | | 鹿眼蛱蝶(Junonia coenia) | 季节性表型可塑性[52] |
| 斑凤蝶(Papilio clytia) | 贝氏拟态[73] |
| 狐眼袖蝶(Heliconius numata) | 米勒拟态[74-75] |
| Distal-less | 转录因子 | 偏瞳蔽眼蝶(Bicyclus anynana) | 眼斑[48] |
未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
mRNA-Seq whole-transcriptome analysis of a single cell
1
2009
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
The molecular basis of scale development highlighted by a single-cell atlas of Bicyclus anynana butterfly pupal forewings
0
2024
Lepidopteran scale cells derive from sensory organ precursors through a canonical lineage
1
2025
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position
1
2013
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
A microRNA is the effector gene of a classic evolutionary hotspot locus
1
2024
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
A long noncoding RNA at the cortex locus controls adaptive coloration in butterflies
1
2024
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
Genome editing retraces the evolution of toxin resistance in the monarch butterfly
1
2019
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...
Adaptive substitutions underlying cardiac glycoside insensitivity in insects exhibit epistasis in vivo
1
2019
... 未来的蝶类研究可在已有研究体系的基础上进一步拓展研究方法和范式,以揭示蝶类适应性演化的深层机制.多组学技术(如转录组学、表观组学、蛋白组学和代谢组学等)的快速发展有助于构建与蝶类发育演化和环境因素响应相关的基因调控网络.单细胞转录组测序[76-78]技术可提高基因表达检测的分辨率,解析细胞类型特异性的表达模式与调控网络.转座酶可及的染色质高通量测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)[79]等实验方法有助于深入挖掘顺式调控元件等非编码序列的功能,以进一步理解复杂表型的遗传机制与演化历史,例如,拟态性状与旱雨季表型可塑性等关键性状的调控[80-81].机器学习与深度学习等技术则可以对蝶类翅型等形态特征进行表型量化,并整合表型-环境-基因表达等多模态信息解析其取食、求偶、迁飞等行为模式.由于蝶类对环境变化高度敏感,已被广泛视作生态环境与生物多样性保护的重要指示物种,在保护生物学中也具有独特价值.在动物资源的仿生利用方面,蝶类研究有助于推动在动物的结构、功能、行为、生理等方面进行仿生利用,例如基于枯叶蛱蝶设计迷彩伪装,具有对环境友好的特征,可推动节能减排和绿色发展.君主斑蝶对强心苷表现出显著的抗性特征,其钠钾离子泵(Na+/K+-ATPase)基因关键氨基酸位点发生了适应性替换[82-83],实现了对毒素的天然耐受.钠钾离子泵作为治疗心力衰竭药物(如地高辛)和多种神经系统疾病的重要作用靶点,在人类生理及病理过程中具有关键功能.蝶类体系中对强心苷抗性的演化机制不仅揭示了物种适应性演化的分子基础,也为深入解析人类相关疾病的生理机制、探索药物靶点选择性调控策略以及耐药性形成机制提供了新的研究视角和思路.因此,蝶类研究体系及实验方法的拓展,将为探索动物适应性演化机制提供新思路,还有望为生态保护、工程应用以及生理疾病等领域提供重要支撑,形成多学科交叉、研究和应用范式. ...










{kind=link}
{kind=link}