








前人研究分析了高原缺氧的发病机制以及代谢组学在高原缺氧相关疾病研究中的应用[5]。本研究基于超高效液相色谱(ultra-high-performance liquid chromatography,UHPLC)-四极杆静电场轨道阱质谱(quadrupole electrostatic field orbital trap-mass spectrometry,QE-MS) 分析方法,鉴定生活于不同海拔的高原鼠兔肾脏差异代谢标志物,探讨标志性差异代谢物及其关键通路,以期为高原鼠兔及高原土著动物肾脏低氧适应机制研究提供实验依据。
1 材料与方法
1.1 实验动物
选取生活于青海省果洛藏族自治州玛多县星宿海地区海拔4 360 m的高原鼠兔(MD组)和藏族自治州门源地区海拔2 900 m的高原鼠兔 (MY组)各 10只,体重均在120~150 g,3月龄左右,雌雄各半。两组高原鼠兔的生存环境均为山地草原,捕捉时间均在11月中旬。MD组动物生存环境温度为-8
1.2 主要试剂和仪器
甲醇(批号167312)、乙腈(批号789101)、乙酸铵(批号112131)、22%氨水(批号145161)和甲酸(批号130050)均为质谱纯,购自德国CNW Technologies公司;L-2-氯苯丙氨酸(批号20240819)购自上海康朗生物科技有限公司。
UHPLC系统(1290 InfinityⅢ LC)购自美国Agilent公司;高分辨质谱仪(Triple TOF 6600)购自美国AB Sciex公司;高速离心机(Heraeus Fresco17)、数据采集和分析软件(Xcalibur 4.0.27)和组合型四极杆静电场轨道阱质谱仪Q Exactive(Orbitrap MS)均购自美国Thermo Fisher Scientific公司;天平(BSA124S-CW)购自德国Sartorius公司;研磨仪(JXFSTPRP-24)购自上海净信科技有限公司;明澈纯水仪(D24 UV)购自德国Merck Millipore公司;超声仪(PS-60AL)购自深圳市雷德邦电子有限公司;ACQUITY UPLC HSS T3色谱柱(2.1 mm×100 mm,1.8 μm)购自美国Waters公司;全自动血液分析仪(ADVIA 2120i)购自德国Siemens公司;全自动生化分析仪(BS-1000M)购自深圳迈瑞生物医疗电子股份有限公司;干式血气分析仪(ST2000)购自武汉明德生物科技股份有限公司。
1.3 血液及肾脏样本采集
所有高原鼠兔在捕捉当天即转运至青海大学医学院/高原医学研究中心的实验室进行采样。按照体重计算麻醉用药剂量,使用8 mL/kg乌拉坦经腹腔注射麻醉后进行腹主动脉采血。将血液常温静置2 h,然后以3 000 r/min、4 ℃离心10 min,收集上清液,分装后置入-80 °C冰箱备用。高原鼠兔的血液样本采集完毕后,注射过量乌拉坦使动物安乐死,然后摘取右侧肾脏,立即置于液氮冷冻,随后转移至-80 °C冰箱保存。本研究中动物实验方案经青海大学医学院医学科学研究伦理委员会审查批准(No. 2021-16),实验操作过程中严格遵循《实验动物管理条例》和《医学实验动物管理实施细则》等规范文件。
1.4 一般生理生化指标测定
采集好的两组高原鼠兔血清样本,一部分经全自动血液分析仪计算红细胞计数、血红蛋白浓度及血细胞比容,另一部分用血气分析仪测定二氧化碳结合力,剩余部分置于冰上用全自动生化分析仪检测葡萄糖含量、尿素氮含量、肌酐含量、同型半胱氨酸含量、尿酸含量等生化指标。
1.5 肾组织代谢组学分析
1.5.1 代谢物提取
称取两组高原鼠兔每只50 mg右肾组织块样品,加入1 000 μL提取液(甲醇、乙腈和水的体积配比为2∶2∶1,且含L-2-氯苯丙氨酸1 μg/mL),涡旋混匀30 s;加入钢珠,45 Hz研磨处理4 min,冰水浴中超声5 min。重复上述步骤2~3次,-20
1.5.2 UHPLC-QE-MS测定及数据处理
使用1290 UHPLC系统进行液相色谱(liquid chromatography,LC)-串联质谱联用(tandem mass spectrometry,MS/MS)分析,使用的UPLC HSS T3色谱柱(2.1 mm×100 mm,1.8 μm )与Q Exactive耦合。每份样品的进样体积为2 μL。流动相A以0.1%甲酸水溶液为正离子模式,以5 mmol/L乙酸铵水溶液为负离子模式;流动相B为乙腈。洗脱梯度设置:0 min,99%A,1%B;1 min,99%A,1%B;8 min,1%A,99%B;10 min,1%A,99%B;10.1 min,99%A,1% B;12 min,99%A,1% B。流速为0.5 mL/min。
在LC/MS实验信息依赖采集(information dependent acquisition,IDA)的基础上,使用Q Exactive质谱仪获取MS/MS数据。Xcalibur 4.0.27软件会根据事先设定的条件,自动采集并触发MS/MS质谱图的生成,同时实时分析全扫描测量的MS数据。同时,电喷雾电离(electrospray ionization,ESI)源参数条件设置:鞘流气(Sheath)气流速为45 Arb,辅助气流(Aux)速为15 Arb,质量即质荷比(m/z)范围为70~1 000,毛细管温度400 ℃,全扫描MS分辨率为70 000,MS/MS分辨率为17 500,噪声对比估计(noise contrastive estimation,NCE)模型中碰撞能量为20/40/60 eV,喷雾电压分别为4.0 kV(正)或-3.6 kV(负)。
使用ProteoWizard软件将质谱原始数据转成mzXML格式。再使用MAPS软件(XCMS内核,1.0版本)做保留时间矫正、峰识别、峰提取、峰积分、峰对齐等预处理,生成由保留时间、m/z值和峰强度组成的数据矩阵。以上UHPLC-QE-MS测定由上海百趣生物医学科技有限公司完成,并利用该公司的内部MS2数据库进行代谢物鉴定。
1.5.3 差异代谢物筛选及信号通路识别
主成分分析(principal component analysis,PCA)通过正交偏最小二乘判别分析(orthogonal partial least squares discriminant analysis,OPLS-DA)过滤代谢物中与分类变量不相关的正交变量,进而发现差异性代谢物。筛选标准为OPLS-DA模型第一主成分的变量投影重要度(variable importance in the projection, VIP)>1.5且倍数变化(fold change,FC)>1.5,或VIP>1.5且FC<1/1.5。对肾脏代谢物样本变量进行PCA、OPLS-DA以及第一、第二主成分的多元变量模式识别分析和OPLS-DA置换检验,并绘制各图。其中,置换检验后用决定系数(R²Y)表示模型对因变量Y 的方差解释程度,即模型能够解释的Y方差的比例;取值范围为 0~1,值越接近 1,表示模型对Y的拟合效果越好。同时用预测决定系数(Q²)评估模型对新数据的预测能力,通常通过交叉验证计算,反映了模型在未参与建模的数据上的预测性能;取值范围为 0~1,值越接近 1,表示模型的预测能力越强。
运用相关性分析热力图、差异显著性分析火山图、信号通路识别气泡图和矩形图分别进行差异代谢物及相关信号通路分析。其中相关性分析热力图中通过颜色深浅表示代谢物之间的相关性强度,颜色越深表示相关性越强;其横轴和纵轴均为代谢物名称,颜色表示相关性系数。火山图以对数变化值为横轴,显著性值为纵轴,通过点的位置和颜色区分显著性差异的代谢物;点的位置越靠近右上角或左上角,表示该代谢物的变化显著且变化幅度大。气泡图的横轴为富集因子(表示代谢物在通路中的富集程度),纵轴为通路名称;气泡大小表示富集的代谢物数量,颜色表示P值的显著性(颜色越深表示P值越小)。利用矩形图进行相关信号通路分析,以通路名称为横轴,富集因子或P值为纵轴,通过矩形的大小或颜色表示通路的富集程度或显著性;矩形越大或颜色越深,表示通路的富集程度越高或显著性越强。
1.6 统计学方法
使用SPSS 27软件进行统计学分析。数据均采用均数±标准差(
2 结果
2.1 不同海拔高原鼠兔的一般生理生化比较
生活于海拔4 360 m和2 900 m的两组高原鼠兔红细胞计数、葡萄糖、尿素氮、肌酐和同型半胱氨酸含量差异无统计学意义(均P>0.05),而血红蛋白浓度、血细胞比容、尿酸和二氧化碳结合力有显著差异(P<0.05,表1),提示生活于不同海拔的高原鼠兔血液中红细胞携氧能力已存在明显差异。其中MD组尿酸含量高于MY组,而血红蛋白、血细胞比容和二氧化碳结合力水平低于MY组,提示随着海拔升高,高原鼠兔对低氧条件出现生理适应,代谢和血液系统产生相应改变。
表1
不同海拔高原鼠兔的一般生理生化指标比较 (
Table 1
项目 Item | MD组 MD group (n=10) | MY组 MY group (n=10) | t值 t value | P 值 P value |
|---|---|---|---|---|
| RBC Ccell/(1012·L-1) | 6.93±0.85 | 6.75±1.65 | 0.355 | 0.726 |
| HGB ρ/(g·L-1) | 116.36±7.39 | 141.40±21.45 | -4.074 | 0.001 |
| HCT φ/(%) | 36.45±2.62 | 42.58±7.09 | -2.985 | 0.007 |
| GLU c/(mmol·L-1) | 6.07±3.66 | 5.84±2.51 | 0.177 | 0.861 |
| BUN c/(mmol·L-1) | 7.48±4.01 | 5.89±3.00 | 1.054 | 0.305 |
| Cr c/(µmol·L-1) | 21.55±5.97 | 22.36±8.65 | -0.258 | 0.799 |
| UA c/(µmol·L-1) | 45.09±21.90 | 24.27±15.84 | 2.555 | 0.019 |
| CO2CP c/(mmol·L-1) | 18.72±5.85 | 26.01±5.83 | -2.928 | 0.008 |
| HCY c/(µmol·L-1) | 25.03±10.31 | 18.35±12.09 | 1.393 | 0.179 |
2.2 不同海拔高原鼠兔的肾脏代谢物出现明显的聚类型分布
PCA、OPLS-DA以及第一、第二主成分的多元变量模式识别分析和OPLS-DA置换检验结果见图1。PCA模型得分散点图(图1A)显示,海拔4 360 m(MD组)和2 900 m(MY组)的高原鼠兔肾脏代谢物样本分散在2个不同的区域;主成分得分值基本集中分布于散点置信区内的区域无明显交叉和重叠,表明两组之间存在肾脏组织主成分代谢物的差异。PCA散点图(图1B)显示,所有样本包括质控用QC样本均基本在95%置信区间(霍特林T²置信椭圆,Hotelling's T-squared ellipse)内。 OPLS-DA模型得分散点图(图1C)显示,MD和MY样品分散在2个不同的区域,呈聚类型分布于圆形散点置信区内的2个区域,结果显示OPLS-DA模式识别效果优于PCA。OPLS-DA模型置换检验(图1D)中R²Y=0.930,Q²=0.655;其中,R2Y非常接近1,说明建立的模型符合样本数据的真实情况;Q2值为0.655,说明模型可以比较好地解释两组样本之间的差异。OPLS-DA模型置换检验(图1D)显示,置换检验随机模型的Q2值均小于原模型的Q2值(0.930 vs 0.840),Q2的回归线与纵轴的截距为-0.61;随着置换保留度逐渐降低,置换的Y变量比例增大,随机模型的Q2逐渐下降,表明建立的模型具有良好的稳健性,不存在过拟合现象。
图1

图1
超高效液相色谱-四极杆静电场轨道阱质谱(UHPLC-QE-MS)分析不同海拔高原鼠兔的肾脏代谢组学
Figure 1
Ultra-high-performance liquid chromatography coupled with quadrupole electrostatic field orbital trap-mass spectrometry (UHPLC-QE-MS) analysis of kidney metabolomics in plateau pikas at different altitudes
2.3 不同海拔高原鼠兔的肾脏代谢物差异
UHPLC-QE-MS分析后,经数据库比对和OPLS-DA筛选标准发现,不同海拔高原鼠兔的肾脏代谢物差异分子有46个,详细数据见表2。其中,海拔4 360 m(MD组)的高原鼠兔肾脏代谢物中蟾蜍二烯羟酸内酯、腺苷、腺嘌呤、薯蓣皂苷、盐酸小檗碱、鼠尾草酚和虾青素等水平明显高于海拔2 900 m的高原鼠兔组(MY组),VIP>1.5,P<0.05;同时,MD组高原鼠兔中花生四烯酸、组胺和香豆素等代谢物的水平低于MY组(VIP>1.5,P<0.05)。
表2 不同海拔高原鼠兔的肾脏代谢物差异分子
Table 2
峰值 Peak | 代谢物 Metabolites | MD组vs MY组 MD group vs MY group | ||||
|---|---|---|---|---|---|---|
| 中文名 Chinese name | 英文名 English name | RT/s | VIP | P value | FC | |
| 4217 | 蟾蜍二烯羟酸内酯 | Bufadienolide | 302.521 | 2.841 | 0.046 1 | 216.417 |
| 3586 | 薯蓣皂苷配基 | Diosgenin | 375.853 | 2.790 | 0.000 0 | 23.644 |
| 2596 | 盐酸小檗碱 | Berberine chloride | 448.647 | 1.684 | 0.010 1 | 2.418 |
| 5668 | 顺丁烯二酰亚胺 | Maleimide | 157.759 | 2.051 | 0.003 3 | 2.275 |
| 6118 | 腺嘌呤 | Adenine | 128.009 | 1.992 | 0.002 6 | 1.355 |
| 4040 | 7,8-脱氢虾青素 | 7,8-Dehydroastaxanthin | 304.975 | 2.124 | 0.007 8 | 13.719 |
| 4168 | 贝那普利酯 | Benazeprilat | 279.705 | 2.212 | 0.000 8 | 10.772 |
| 4196 | 胆固醇-3,7,12,25-呋喃-3-葡萄糖醛酸 | Cholestane-3,7,12,25-tetrol-3-glucuronide | 273.513 | 2.149 | 0.001 2 | 5.633 |
| 4221 | 7-乙氧基香豆素 | 7-Ethoxycoumarin | 261.173 | 1.795 | 0.017 9 | 2.839 |
| 3212 | 1-硬脂酰甘油磷酸甘油 | 1-Stearoylglycerophosphoglycerol | 412.530 | 1.586 | 0.025 1 | 2.103 |
| 5578 | 2-甲氧基对乙酰氨基酚硫酸盐 | 2-Methoxyacetaminophen sulfate | 36.282 | 2.150 | 0.000 5 | 0.453 |
| 5294 | 黄嘌呤 | Xanthosine | 57.261 | 1.863 | 0.007 7 | 0.448 |
| 5476 | 四氢生物蝶呤 | Tetrahydrobiopterin | 42.319 | 1.632 | 0.011 0 | 0.425 |
| 2896 | 小穗苎麻素 | Cryptopleurine | 440.968 | 2.258 | 0.000 3 | 0.292 |
| 3198 | 坎二酸 | Canbidiolic acid | 413.844 | 2.004 | 0.012 7 | 0.284 |
| 5570 | GDP-3,6-二脱氧-D-半乳糖 | GDP-3,6-dideoxy-D-galactose | 36.317 | 1.735 | 0.001 6 | 0.197 |
| 5501 | 棉酚 | Gossypol | 41.544 | 1.725 | 0.019 3 | 0.160 |
| 4549 | 4-香豆素基莽草酸酯 | 4-Coumaroylshikimate | 179.435 | 2.266 | 0.009 7 | 0.103 |
| 4596 | 紫丁香苷 | Syringin | 169.451 | 2.374 | 0.010 3 | 0.050 |
| 5805 | 5-O-甲基-间氨基-肌醇 | 5-O-Methyl-myo-inositol | 33.358 | 2.543 | 0.000 5 | 0.026 |
| 5001 | 癸二酸 | Sebacic acid | 135.122 | 2.295 | 0.046 3 | 0.077 |
| 4671 | 萨马素A | Samaderin A | 163.209 | 1.535 | 0.037 8 | 0.217 |
| 4717 | 3-脱氢酶 | 3-Dehydrogenase | 157.505 | 2.158 | 0.000 9 | 0.170 |
| 4514 | 硫烯草丹 | Sulfallate | 185.215 | 1.844 | 0.049 0 | 0.190 |
| 5688 | 双甲苯苄醇 | Bitolterol | 35.519 | 1.568 | 0.001 1 | 0.271 |
| 5523 | 烟碱 | Nicotiamine | 169.858 | 2.201 | 0.000 1 | 0.454 |
| 7265 | 吡达隆 | Pyridarone | 29.322 | 1.490 | 0.019 5 | 0.401 |
| 3058 | 高肌肽 | Homocarnosine | 412.238 | 1.966 | 0.006 3 | 0.390 |
| 3410 | 阿非迪霉素 | Aphidicolin | 388.298 | 2.133 | 0.005 0 | 0.205 |
| 3101 | 松香酸盐 | Abietate | 408.903 | 2.373 | 0.033 4 | 0.088 |
| 4573 | 秦皮苷 | Fraxin | 174.802 | 1.098 | 0.034 3 | 0.085 |
| 3554 | 高多环内酯 | Homodolicholide | 380.038 | 1.667 | 0.004 0 | 0.234 |
| 3984 | 法尼基半胱氨酸 | Farnesylcysteine | 328.343 | 2.148 | 0.015 9 | 0.224 |
| 59 | 泛酸 | Pantothenic acid | 157.754 | 2.460 | 0.000 1 | 0.480 |
| 6640 | 1-甲基腺苷 | 1-Methyladenosine | 44.286 | 2.135 | 0.003 3 | 0.482 |
| 5674 | L-二氢脱壳素 | L-Dihydroanticapsin | 157.754 | 2.510 | 0.000 1 | 0.463 |
| 3695 | 十八碳四烯酸 | Stearidonic acid | 359.953 | 2.698 | 0.003 7 | 0.103 |
| 4631 | 6-脱氧巴西红厚壳素 | 6-Deoxyjacareubin | 237.103 | 2.310 | 0.011 0 | 0.096 |
| 5940 | 异戊烯焦磷酸 | Isopentenyl diphosphate | 142.170 | 2.771 | 0.038 3 | 0.004 |
| 6407 | 环氧化物青蒿琥酯 | Artomunoxanthentrione epoxide | 27.729 | 2.401 | 0.000 0 | 0.249 |
| 5768 | 苄基青霉酸 | Benzylpenicilloic acid | 151.870 | 2.655 | 0.025 4 | 0.034 |
| 302 | 异亮氨酸 | Isoleucine | 71.954 | 1.669 | 0.045 9 | 0.698 |
| 6496 | L-缬氨酸 | L-Valine | 55.803 | 1.684 | 0.018 2 | 0.637 |
| 4165 | 鼠尾草酚 | Carnosol | 279.762 | 2.070 | 0.003 8 | 31.391 |
| 41 | 花生四烯酸 | Arachidonic acid | 466.526 | 1.614 | 0.010 5 | 0.695 |
| 5711 | 组胺 | Histamine | 35.217 | 1.740 | 0.042 9 | 0.537 |
2.4 不同海拔高原鼠兔肾脏差异代谢物的相关性及信号通路
对不同海拔高原鼠兔肾脏差异代谢物的定量值进行Pearson相关系数计算,结果以热力图表现(图2A)。图中,横纵坐标代表两组对比的差异代谢物,不同位置的色块代表对应位置代谢物间的相关系数大小,红色表示正相关,蓝色表示负相关;非显著性相关用叉号进行标注。正相关r值为0~1,负相关r值为-1~0;r的绝对值越接近1,两变量的关联程度越强;反之,r绝对值越接近0,关联程度越弱。
图2
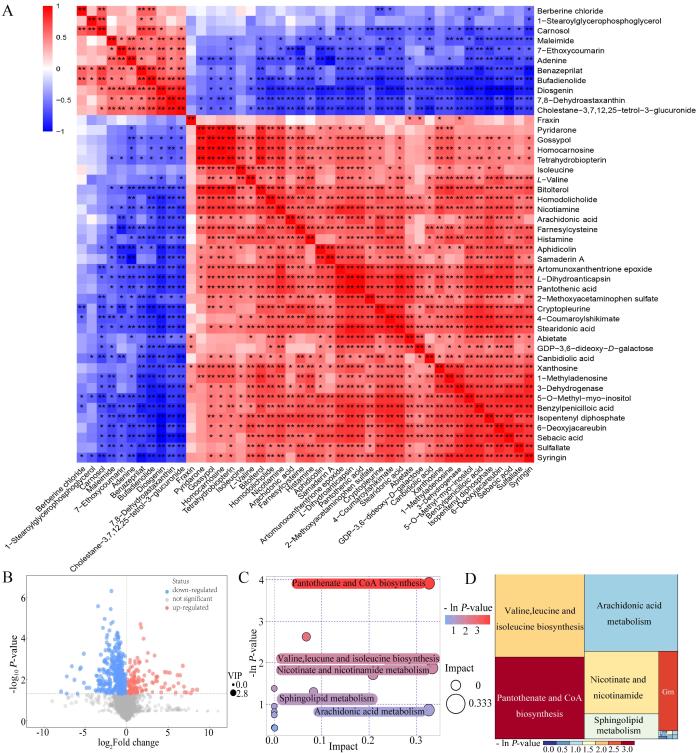
图2
不同海拔高原鼠兔的肾脏差异代谢物相关性及其相关通路
Figure 2
Correlation of kidney differential metabolites and related pathways in plateau pikas at different altitudes
火山图(图2B)中,每个点代表一个代谢物,横坐标代表该组对比各物质的FC(取以2为底的对数),纵坐标表示t检验的P值(取以10为底的对数的负数),散点大小代表OPLS-DA模型的VIP值(即散点越大,VIP值越大);散点颜色代表最终的筛选结果,显著上调的代谢物以红色表示,显著下调的代谢物以蓝色表示,非显著差异的代谢物以灰色表示。
3 讨 论
青藏高原有着独特的地理环境和气候特征。高原鼠兔是青藏高原土著小型啮齿类动物,已经进化出特有的遗传和代谢机制来适应严峻的高原环境。有文献报告,高原低氧暴露会造成不同程度的肾功能损伤[6]。然而高原低氧环境下高原土著动物肾脏适应的代谢组学研究报告甚少。本研究选取生活于4 360 m和2 900 m海拔的高原鼠兔,鉴定其肾脏差异代谢物,并探讨其可能的低氧适应机制。通过OPLS-DA建模分析显示,该模型符合样本数据的真实情况,具有高解释力和稳健性;PCA模型的散点图也表明两组样本间存在明显差异。结果表明,两组高原鼠兔肾脏在适应高原低氧等特殊环境时存在不同的代谢物质基础。
由于高海拔氧分压的降低,高原鼠兔促红细胞生成素刺激红细胞增殖和血红蛋白浓度升高,从而增强血液携氧能力,确保组织能够获得足够的氧气。此外,高海拔高原鼠兔的尿酸水平升高可能与慢性缺氧环境下红细胞增多相关,血红蛋白嘌呤代谢产物尿酸也随之增多。尿酸在血清中具有抗氧化作用,能保护红细胞免受氧化应激损伤,维持红细胞形态,阻止棘形和球形红细胞的形成。高海拔地区的氧分压下降,动物需要增加呼吸频率和呼吸深度来补偿氧气的不足,这会导致呼出的二氧化碳量增加,二氧化碳结合力下降。
高原鼠兔受到长期低氧胁迫表现出适应高原环境的遗传和代谢机制改变。本研究表明,生活于海拔4 360 m的MD组高原鼠兔肾脏组织中腺苷水平均显著大于生活于海拔2 900 m的MY组。腺苷有助于血管扩张和拮抗儿茶酚胺类物质的缩血管效应。有文献报告,在极端高海拔环境条件下,细胞外腺苷水平也随之升高[7]。然而腺苷二磷酸(adenosine diphosphate,ADP)和腺苷三磷酸(adenosine triphosphate,ATP)有助于细胞外腺苷的生成;在疾病状态下,外核苷酸酶表达增加和腺苷生成是保护细胞和抑制炎症的机制之一[8]。腺嘌呤经补救合成途径生成腺苷一磷酸(adenosine monophosphate,AMP)[9]。由此推测,MD组高原鼠兔的腺嘌呤增加可能是机体能量利用的适应机制之一。
有研究发现,鼠尾草酸和鼠尾草酚可激活Kelch样ECH相关蛋白1(Kelch-like ECH-associating protein 1,Keap1)/核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,Nrf2)/抗氧化反应元件(antioxidant response element,ARE)信号转导通路而发挥抗氧化作用[14]。因此推测,低氧刺激导致高原鼠兔体内氧化应激增加,Keap1/Nrf2/ARE经典抗氧化应激通路激活,这可能是本研究中MD组高原鼠兔的鼠尾草酚水平显著高于MY组的分子机制。另外有文献报道,虾青素可激活Nrf2,促进Keap1与Nrf2解离,进而促进ARE下游基因氧化还原平衡因子表达,有利于高原鼠兔适应高原低氧条件下氧化应激环境。Hassanein 等[15]研究发现,17种植物来源的香豆素衍生物可刺激Nrf2转录,致使下游Keap1/ Nrf2/ARE通路激活,从而发挥抗氧化应激损伤效应。组胺对肾小球滤过率的影响被认为是组胺与H1受体介导肾小动脉收缩调节的结果[16-17]。本研究中,MD组的组胺水平显著低于MY组,推测低氧环境激活高原鼠兔肾脏组织中组胺H1受体激活,导致入球动脉收缩,肾小球血浆流量减少,肾小球滤过率下降。
本研究中氨基酸代谢差异表明其在肾脏高原适应机制中发挥核心作用,它们不仅是蛋白质合成的基本单元,还参与肾脏的多种代谢过程。肾脏中氨基酸通过不同的转运系统被重吸收和分泌,维持血液中氨基酸的平衡,并支持肾脏在滤过、重吸收和排泄代谢废物方面的功能。泛酸盐(维生素B5)及其衍生物辅酶A在肾脏适应中扮演重要角色。泛酸盐是辅酶A的前体,而辅酶A是脂肪酸氧化、糖异生和乙酰化反应等代谢通路的关键辅助因子。辅酶A有助于调节能量代谢与脂质代谢,确保肾脏适应高原低氧下的代谢改变,同时肾脏中氨基酸代谢改变对于维持内环境稳态至关重要。氨基酸代谢差异改变可作为高原鼠兔适应高原低氧的特征性改变,对于不同海拔高原鼠兔的适应机制研究具有重要意义。
支链氨基酸(branched-chain amino acids,BCAAs)包括缬氨酸、亮氨酸和异亮氨酸,在蛋白质合成、能量产生和信号转导中起着重要作用。它们的生物合成途径主要涉及丙酮酸衍生物类的氨基酸合成,包括丙酮酸脱羧过程中,丙酮酸与硫胺素焦磷酸(thiamine pyrophosphate,TPP)结合形成中间产物乙酰TPP,再与另一个丙酮酸缩合,形成α-乙酰乳酸,然后通过一系列反应生成BCAAs。BCAAs的代谢与蛋白质合成和细胞生长密切相关,其作用机制主要通过mTOR信号通路实现。泛酸是辅酶A的前体,二者均为生物体内重要的代谢物,在能量代谢及生物合成中发挥关键作用。辅酶A参与如脂肪酸氧化、糖酵解及三羧酸循环等生化反应。
蟾蜍二烯羟酸内酯、腺苷、腺嘌呤、薯蓣皂苷、盐酸小檗碱、鼠尾草酚和虾青素等代谢物,以及花生四烯酸、组胺和香豆素等,在生物体内的代谢过程可能与BCAAs和辅酶A的生物合成途径存在交互影响。例如,氨基酸代谢可能影响上述代谢物生成,而辅酶A作为酰基载体参与脂肪酸的合成和氧化,也可能影响上述代谢物的生成。上述代谢物的生物合成和代谢途径可能受到BCAAs和辅酶A代谢的影响,因为其可能共享相同的代谢中间产物或由共同信号通路调控。总而言之,BCAAs和辅酶A的生物合成途径与上述代谢物之间存在复杂的交互调控,对于维持生物体的正常代谢过程发挥着重要作用。本研究中,缬氨酸、亮氨酸和异亮氨酸生物合成通路及泛酸和辅酶A的生物合成通路的影响因子和富集程度依次下降且具有显著性,表明上述通路可能在高原鼠兔低氧适应方面具有重要作用。本研究还发现其他一些高VIP值、P值和FC值的差异性代谢物,如环氧杂蒽酮在MD组表达水平显著低于MY组,而藜芦胺和顺丁烯二酰亚胺等表达水平在MD组显著高于MY组。这些差异代谢物在低氧环境的调节机制目前仍不明确,还需进一步研究。
综上所述,本研究中发现的肾脏组织差异代谢物可能揭示不同海拔高原鼠兔的高原适应机制,可作为低氧适应的生物标志物,并为高海拔地区人类高原肾脏相关疾病治疗提供新的研究方向。
[引用本文]
何雨昕, 白振忠, 薛华, 等. 基于UHPLC-QE-MS的高原鼠兔肾脏差异代谢物及低氧适应机制分析[J]. 实验动物与比较医学, 2025, 45(1): 3-12. DOI: 10.12300/j.issn.1674-5817.2024.095.
HE Y X, BAI Z Z, XUE H, et al. Analysis of kidney differential metabolites and hypoxia adaptation mechanism of plateau pikas based on UHPLC-QE-MS[J]. Lab Anim Comp Med, 2025, 45(1): 3-12. DOI: 10.12300/j.issn.1674-5817.2024.095.
医学伦理声明
本研究方案经青海大学医学院医学科学研究伦理委员会审查批准(No. 2021-16)。所有实验过程均遵照中国实验动物相关法律法规条例要求进行。
Medical Ethics Statement
This study has been reviewed and approved by the Medical Research Ethics Committee of Qinghai University Medical School (No. 2021-16). All experimental operations have been carried out in accordance with the requirements of the relevant laws and regulations on experimental animals in China.
作者贡献声明
何雨昕负责血液生化分析、质谱分析、信号通路等生物信息学分析,制图制表,整理数据,统计分析,撰写初稿;
白振忠负责动物捕捉,参与研究设计;
薛华参与动物捕捉、采样、血液生化分析、质谱分析、信号通路等生物信息学分析,以及制图制表;
郭子旭参与动物捕捉、采样、血液生化分析、质谱分析,以及信号通路等生物信息学分析;
曹学锋负责动物捕捉,研究设计和修订。
利益冲突声明
所有作者均声明本文不存在利益冲突。
参考文献
高原鼢鼠和高原鼠兔血液参数和血红蛋白亚型对不同海拔生境的响应
[J].
Responses of blood parameters and hemoglobin subtypes in plateau zokors and plateau pikas to different altitude habitats
[J].
高原低压低氧环境对平原健康男青年血像和肾功能影响
[J].
Impact of hypoxia environment at plateau on renal function among healthy male youth
[J].
低氧暴露下高原鼠兔血液中三种内源性气体分子含量的变化
[J].
Changes of three endogenous gas molecules in the blood of plateau pika exposed to hypoxia
[J].
低氧暴露下高原鼠兔肺组织间隙连接蛋白40表达分析
[J].
Expression analysis of gap junction protein 40 in lung of plateau pika exposed to hypoxia
[J].
Metabonomics window into plateau hypoxia
[J].
急性肾功能损伤在急性高原病发病过程中的差异研究
[J].
Phenotypes of acute renal function injuries in different acute high-altitude diseases
[J].
Pharmacology of adenosine receptors: the state of the art
[J].
Adenosine at the interphase of hypoxia and inflammation in lung injury
[J].
Increased synaptic ATP release and CD73-mediated formation of extracellular adenosine in the control of behavioral and electrophysiological modifications caused by chronic stress
[J].
Diosgenin: an updated pharmacological review and therapeutic perspectives
[J].
Recent advances in the pharmacological activities of dioscin
[J].
Biological activity of berberine-a summary update
[J].
Neural control of blood pressure in chronic intermittent hypoxia
[J].
Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs
[J].
Coumarins as modulators of the Keap1/Nrf2/ARE signaling pathway
[J].
The implications of histamine metabolism and signaling in renal function
[J].
Histamine in the kidneys: what is its role in renal pathophysiology?
[J].
蟾皮化学成分和药理活性的研究进展
[J].
Research progress on chemical constituents and pharmacological activities of Bofonis Corium
[J].
Pathogenesis of pre-eclampsia: marinobufagenin and angiogenic imbalance as biomarkers of the syndrome
[J].
New insights on the role of marinobufagenin from bench to bedside in cardiovascular and kidney diseases
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}